Y-DNA ハプログループと
日本人の起源
作成:2025/01/20
追記:2025/07/06
- 目次
Y-DNA ハプログループの分岐
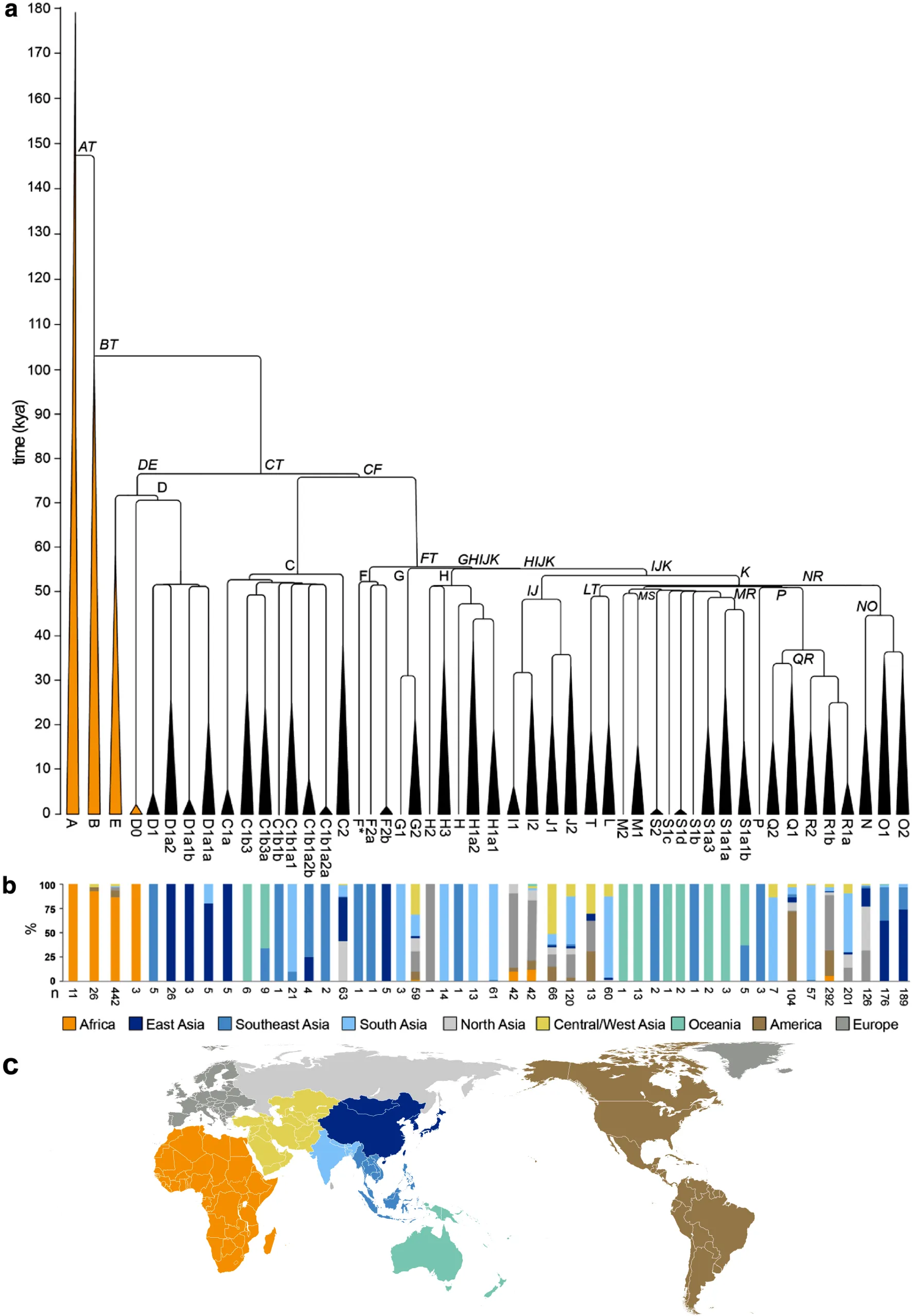
下はウィキペディアの「Y-DNAハプログループ」の項目に 2025/01/20 現在掲載されている Y 染色体ハプログループの系統と分岐年を示す図(ハラスト氏らによる)である。
系統と分岐年
- 系統図をクリック(タップ)すると拡大
出アフリカ組(D1, C, FT)の下位系統の放散は約5万年前に集中している。PからRとQへの分岐は例外的に約3万5000年前に起きている。(=系統樹作者のコメント)
出典(開|閉)
Pille Hallast, Anastasia Agdzhoyan, Oleg Balanovsky, Yali Xue & Chris Tyler-Smith - Hallast, P., Agdzhoyan, A., Balanovsky, O. et al. A Southeast Asian origin for present-day non-African human Y chromosomes. Hum Genet (2020). https://doi.org/10.1007/s00439-020-02204-9, CC 表示 4.0, リンクによるこの系統樹によると、
- A 系統から AT 系統の分岐が約147,000年前。
- AT 系統から B 系統と CT 系統に分岐したのが約 103,000 年前。
- CT 系統から DE 系統と CF 系統に分岐したのが約 77,000 年前。
- DE 系統から D 系統と E 系統が分岐したのが約 72,000 年前。
- D 系統から D0 と D(0以外)が分岐したのが約 70,000 年前。
- D(0以外) 系統から(D1 + D1a2)、(D1a1a + D1a1b)が分岐したのが約 50,000 年前。この2つはそれぞれすぐに分岐。
-
- D1
- D1a2(縄文系)
- D1a1(チベット民族に多い)
-
- D(0以外) 系統から(D1 + D1a2)、(D1a1a + D1a1b)が分岐したのが約 50,000 年前。この2つはそれぞれすぐに分岐。
- D 系統から D0 と D(0以外)が分岐したのが約 70,000 年前。
- CF 系統は約 75,000 年前に C 系統と FT 系統に分岐。以下別記
- DE 系統から D 系統と E 系統が分岐したのが約 72,000 年前。
- CT 系統から DE 系統と CF 系統に分岐したのが約 77,000 年前。
- AT 系統から B 系統と CT 系統に分岐したのが約 103,000 年前。
- CF 系統は約 75,000 年前に C 系統と FT 系統に分岐。
- C 系統は約 53,000 年前に C1 と C2 に分岐。C1 は約 47,000 年前までに細かく分岐。
- C1
- C1a(C1a1 は日本人に数 % 存在)
- C2(モンゴル人など遊牧民族に多いが、ネイティブ・アメリカンのナデネ語族にも認められ、日本人にも地域差が大きいものの数 % 存在)
- C1
- FT 系統は約 55,000 年前に F 系統と GHIJK 系統に分岐。
- GHIJK 系統はすぐに G 系統と HIJK 系統に分岐。
- G 系統は約 30,000 年前になってから G1 と G2 に分岐。
- HIJK 系統はすぐに H 系統と IJK 系統に分岐。
- H 系統は約 50,000 年前に(H + H1)と(H2 + H3)に分岐
- IJK 系統は約 52,000 年前に IJ 系統と K 系統に分岐。以下別記
- GHIJK 系統はすぐに G 系統と HIJK 系統に分岐。
- C 系統は約 53,000 年前に C1 と C2 に分岐。C1 は約 47,000 年前までに細かく分岐。
- IJK 系統は約 52,000 年前に IJ 系統と K 系統に分岐。
- IJ 系統は約 47,000 年前に I 系統と J 系統に分岐。
- K 系統は約 50,000 年前に LT 系統、MR 系統に分岐。
系統は変異の蓄積なので、分岐の順番は問題ないであろうが、年代がこの通りであるかどうかは更に検証が必要である。
連結すると下の図のようになった。
なお、連結するにあたって、
- 日本人の起源に近いアルファベット1文字の系統を赤枠で囲んだ
- 日本人に特徴的な系統にラベンダー色の背景色を付けた
- 日本人の起源にほとんど関係ない系統は初期状態では閉じた(必要に応じて開閉可能)
- ウィキペディアの Y-DNA 関係各ページを参考にした
と、日本人の起源関連に特化した。分岐年代は省いた。
- A
- 横スクロール可能
日本人に特徴的なY染色体ハプログループ
- ウィキペディアの「Y-DNAハプログループ」の項目から日本人関連の部分を抜粋。ただし O 系統のハプログループ4つは O 系統としてまとめた。
- D1a2 主に日本で検出され、アイヌと沖縄本島南部(島尻)に特に多い。日本国外では、韓国で平均2%ほどの男性に見られるほか、ミクロネシア、ティモール島、中国(東北部、北京等)でも散発的に検出された例がある。縄文人に由来すると推定される。
➥ウィキペディア「ハプログループD (Y染色体)」 - C1a1 日本固有。現代日本人の5%~6%ほどの男性に見られる。
➥ウィキペディア「ハプログループC1a1 (Y染色体)」 - N1a2 ウラル山脈以東、ネネツ人などのサモエード語派と関連。現代の中国、ベトナム、北朝鮮・韓国、日本などでも低頻度に見られるが、古代人骨から得られたデータを考慮すれば新石器時代から青銅器時代にかけて中国東部(山東省)及び北部(河北省[3]、遼寧省、内蒙古自治区)に於いて主要なハプログループであった模様。
➥ウィキペディア「ハプログループN (Y染色体)」 - O 系統
➥ウィキペディア「ハプログループO (Y染色体)」- O1a 中国東南部、台湾島(とりわけ台湾原住民)、海域東南アジア(フィリピン、マレーシア、インドネシア、シンガポール等)で多く、アドミラルティ諸島やマジュロなどでも10~30%程見られる。オーストロネシア語族と関連。インドシナ半島、中国のその他の地域、韓国、日本、北アジア、中央アジアでも低い頻度に見られる(英語版)。
- O1b1 中国南部、インドシナ半島、インドネシア、インド北東部、ニコバル人、ションペン人、ムンダ族に特に多い。オーストロアジア語族との関連が想定されているが、漢族や日本人に比較的多く見られる下位系統も現存している。
- O1b2 日本人、朝鮮民族に多く(約30%)、満州族、ナナイ、ダウール族、蒙古族、漢族等にも少数見られる。ベトナムやミクロネシアで散発的に観察された例もある。下本山岩陰遺跡や青谷上寺地遺跡で発見された弥生時代の人骨から検出されたことがある。O1b1 と O1b2 の最も近い共通祖先は三万年以上前にも遡ると推定されている。
- O2 中国大陸、朝鮮半島、台湾島、漢民族、朝鮮民族、タイ人、ビルマ族、ヤオ族、シェ族、キン族に多い。日本人にも15%~20%程見られる。シナ・チベット語族、ミャオ・ヤオ語族と関連。石川県金沢市観法寺町にある岩出横穴墓から出土した古墳人一体の遺骨に見られたハプログループ O2a2b1a1a1a4a1-CTS5308 は東アジア全体で比較的多く見られる O-M117 のサブクレードではあるが、O-CTS5308 は特にチベット人に多く見られるタイプである。
- 日本人起源関連 Y 染色体ハプログループの分布
-
ハプログループ D の分布 .png)
- YDNAmapa, CC BY-SA 4.0, via Wikimedia Commons
ハプログループ O の分布 _edit.png)
- Maulucioni, CC BY 3.0, via Wikimedia Commons
ハプログループ N の分布 .png)
- Maulucioni, CC BY 3.0, via Wikimedia Commons
ハプログループ C2(旧C3) の分布 .png)
- Maulucioni, CC BY-SA 3.0, via Wikimedia Commons
.png){kind=link}
.PNG){kind=link}
.PNG){kind=link}
.PNG){kind=link}
なお、日本人祖先には直接関係ないが、謎のハプログループが2つ存在するらしい。
ひとつは、日本人に数 % 存在する C1a1 の最近縁 C1a2 である。ウィキペディアの「ハプログループ C1a1(Y染色体)」によると、35,000 年前前後のヨーロッパ、約 7,000 年前のスペイン北西部とハンガリー、現代ヨーロッパ人、カビル人、アルメニア人、ネパール人にも検出されるとのこと。
もうひとつの謎のハプログループは R 系統である。R1a は中央ヨーロッパからインド、中央シベリアを中心に、飛び地としてシベリア北東端に分布する。R1b は西ヨーロッパから中央アジアを中心に分布し、東シベリアにも稀薄に分布する。分布パターンは N 系統に似ているが、N 系統の方が全体的に北東よりであり、中国東部にも分布する。
Y-DNA ハプログループの拡散ルート
上記のデータから拡散ルートを考えるのが難問である。
従来は、2005~2011年に精力的に論文や著作を執筆した 崎谷満氏の説に基づいて、北ルート(草原ルート?)が重視された拡散が想定されていた。しかし、当然ながらこの十数年間の進展やデータ蓄積が反映されていない。
これに対して、2020年に発表されたハラストらの説では、
- 出アフリカ系統は、速やかにユーラシア全体にわたって拡散し、後に東南アジアで多様な系統に分岐した
- 約55,000-50,000年前に東南アジアから西に逆流した系統は、拡散先住民のY染色体系統と置き換わった
ことが主張されている。
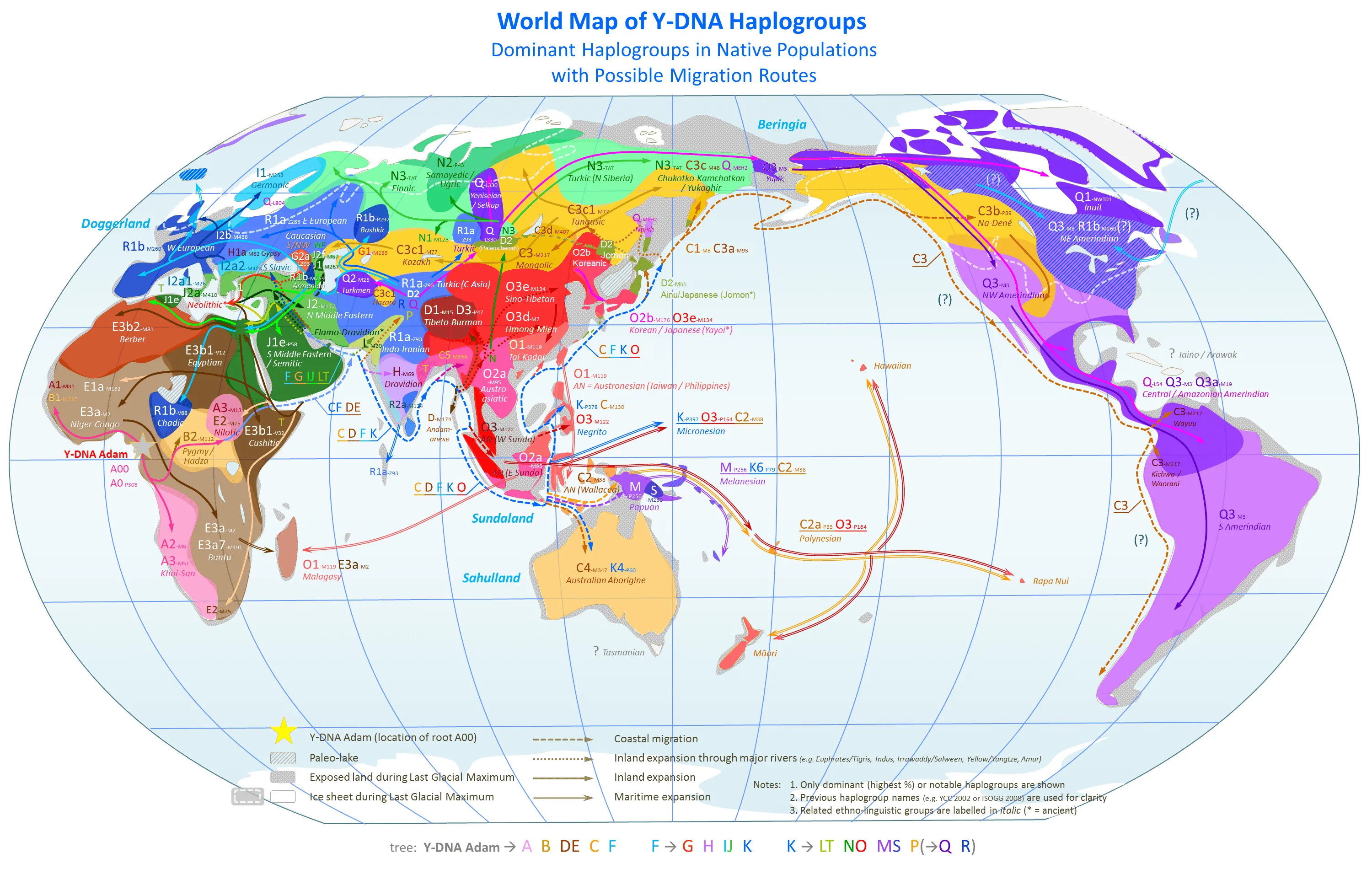
ウィキペディアの「Y-DNAハプログループ」の項目には、2025年1月現在、下の想定地図が掲載されている。
世界拡散を表す想定地図
- 地図をクリック(タップ)すると拡大
- Chakazul, CC BY-SA 3.0, via Wikimedia Commons
{kind=link}
上の地図は、ハプログループの命名方法が最新ではないものの、崎谷氏の説よりも東南アジア経由の Y-DNA 拡散が強調され、逆に北回り(草原の道)経由のルートがトーン・ダウンされている。崎谷氏の説とハラスト氏の折衷になっているようである。
C 系統と O 系統が北回り(草原の道)ではなく、ミャンマーから東進したように矢印が描かれている点で、崎谷氏の説よりも現実的である。しかし、D 系統がミャンマーからチベット、トゥルファン盆地、モンゴル高原、沿海州を経てはるばる日本列島に流入したという考えは不自然に思える。極大期でないにしても、氷河期にチベットを縦断するのは、険阻な地形、酷寒の気候、食糧と水の不足......と困難すぎるのではないだろうか? アフガニスタンとイランの国境付近の高原を通って中央アジアに抜けた方がまだマシである。しかも現代でも同じであるが、富士山よりも標高が高いため空気が非常に薄い。ミャンマーから東進したり、スンダランド経由の方が容易だと思われる。
また、残念ながら制作者は、最終氷期における日本列島周辺の古地理には詳しくないようである。想定地図を拡大すればわかるが、少なくとも、
- 日本海が湖として扱われている
- 沖縄トラフが無視されている
- 千島列島とアリューシャン列島の島々の多くが無視されている
と不適切な点がある。(※地図「最終氷期極大期の北太平洋」)
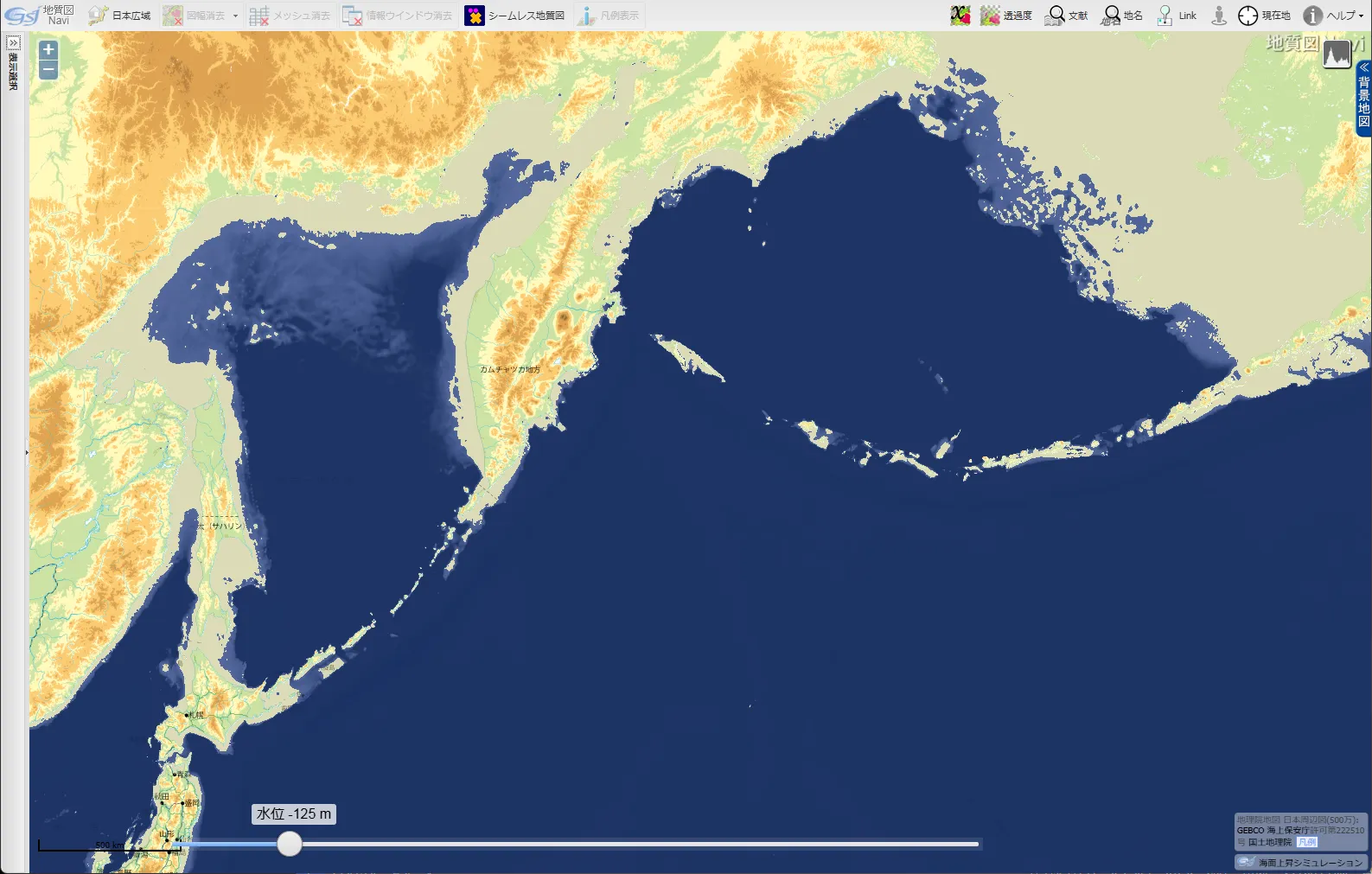
- クリック(タップ)すると拡大
オホーツク海も中央部しか海として描かれていない。ただしこれは流氷が現代よりもずっと大規模だったということを表しているのかも知れない。
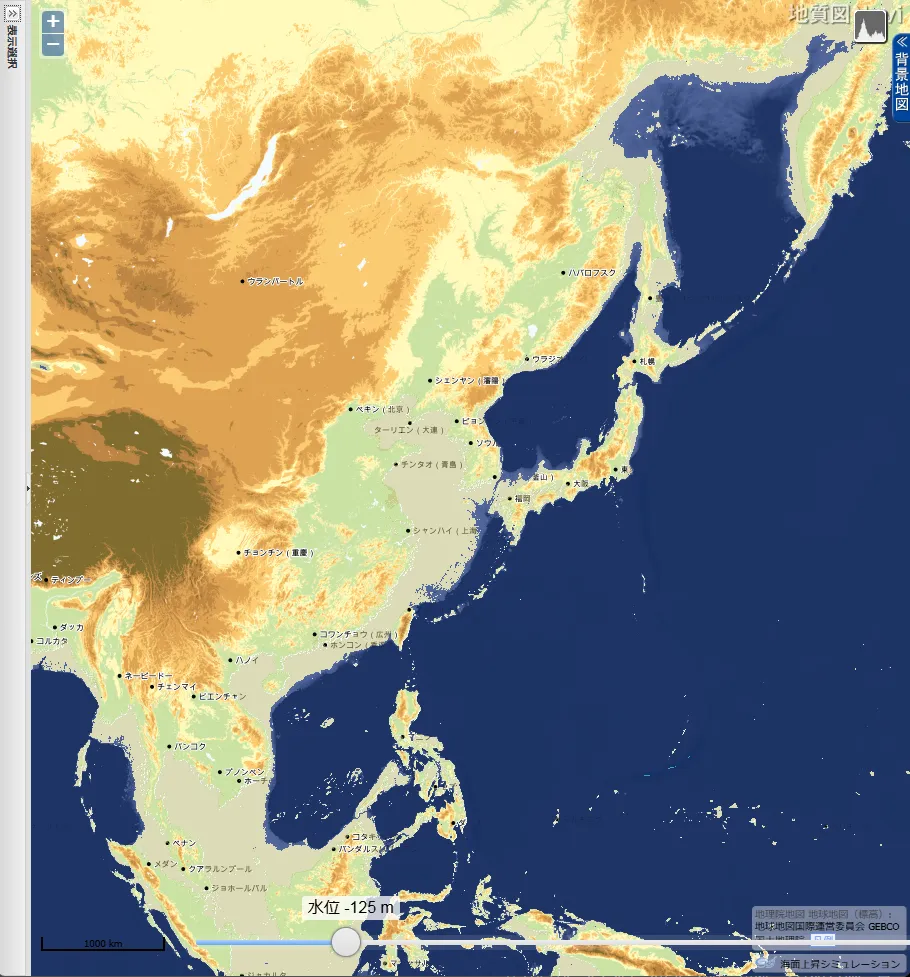
最終氷期における日本列島周辺の古地理を知るのに大いに役立つのが次の章で採り上げる産業総合研究所のサイト「地質図 Navi」の「海面上昇シミュレーション」であり、欲を言えばいくつかの局面に分けてもらいたいところである。
結局「Y染色体ハプログループの世界拡散を表す想定地図」は、現代のY染色体ハプログループ分布地図に最終氷期の陸化していた大陸棚を追加したもののように思える。1枚の地図だけで数万年のY染色体ハプログループの移動を表現するには無理があるので、次の時期の地図が必要であろう。
- 寒冷化停滞期(約50,000~33,000年前)
- 寒冷化・海水位低下進行期(約33,000~25,000年前)
- 最終氷期極大期(約25,000~16,000年前)
- 温暖化・海水位上昇進行期(約16,000~11,500年前)――寒の戻りである12,900年前~11,500年前のヤンガードリアス期を含む。
- 海水位継続上昇期(約11,500~8,000年前)
- 海水位安定期(約8,000年前以降)――約7,000年前~5,000年前のいわゆる「気候最温暖期}(≒縄文海進期)を含む
2025/01/10 現在、一般人に入手しやすい最終氷期後の海水位上昇のグラフとして、ウィキペディアの「海水位変動」の項目に下のグラフが掲載されている(このサイトの別ページでも引用している)。
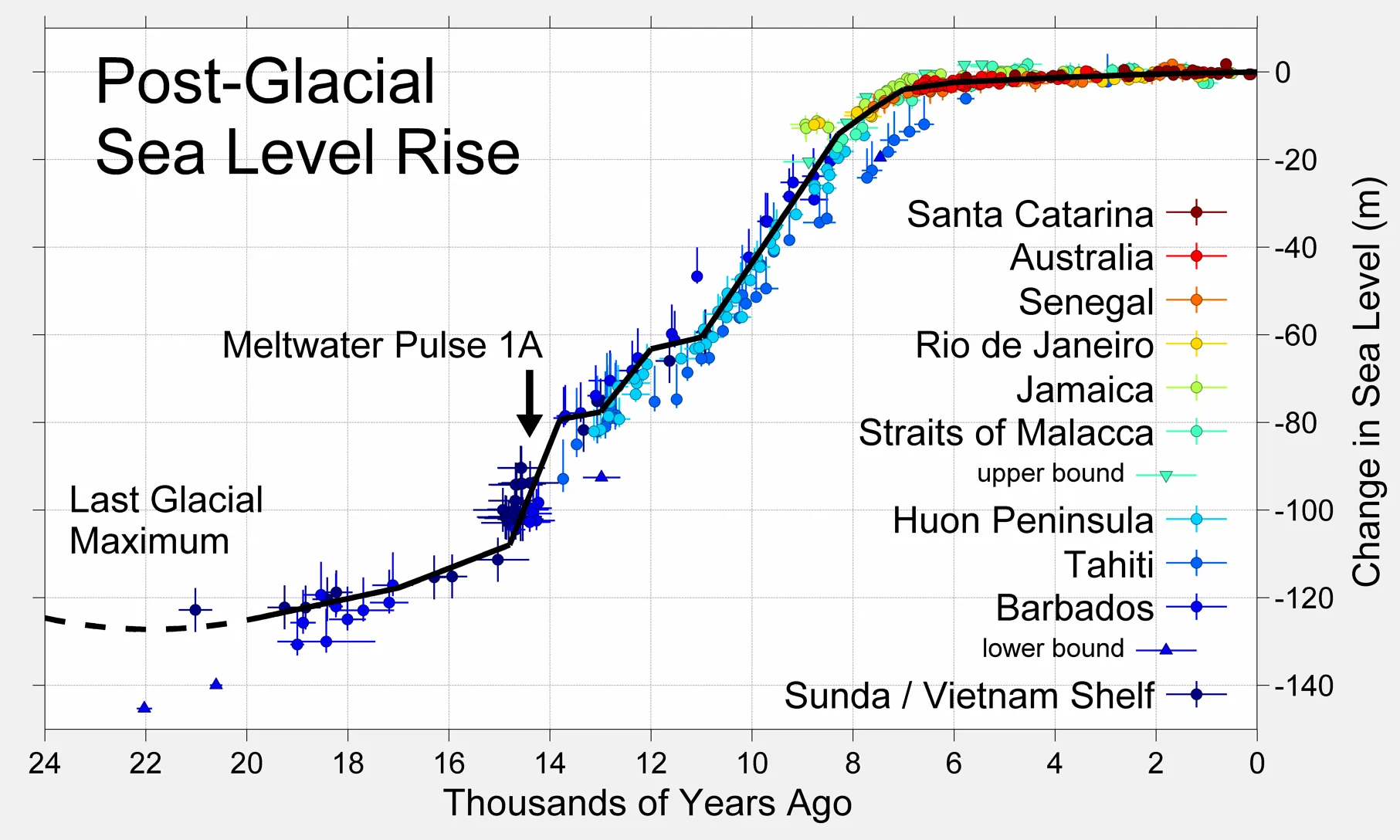
- クリックすると拡大
- 画像出典:Global Warming Art
しかしこのグラフには、元データがどこにあって、どうやってこのグラフになったかという経緯が明確でないため取り下げるべきであるという意見が付記されている。また、出典元となるサイトが売りに出されているのも不審である。
そこで、日本古代史ネットワークの丸地三郎氏が勉強会のためにいくつかの資料を合成し、コメントを加えたグラフを下に掲載する。
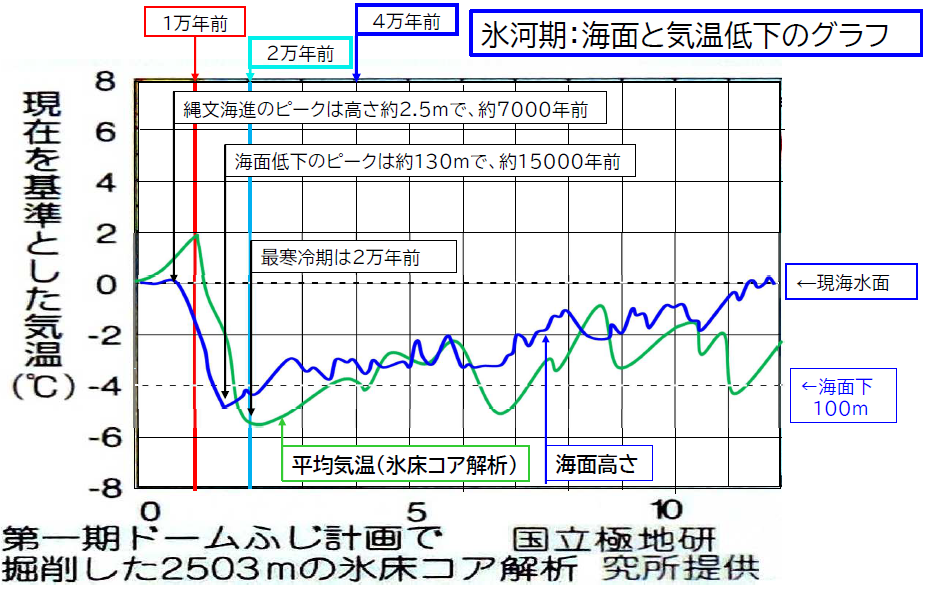
- 元データ(青線グラフ):
- 小野有吾ほか「北海道の自然史-氷期の森林を旅する」(北大図書刊行会)の図
- 田辺晋ほか「東京低地臨海部の沖積層に見られる湾口砂洲の形成」の図
- 上記データを丸地三郎氏が組み合わせ、海面低下のグラフとして合成。
丸地氏が合成したグラフでは、平均気温は南極の氷床コアから解析された値が採用されているが、
- 海水位低下は寒さに約2,000年遅れる。LGM の寒さのピークは約20,000年前(現在よりも約5.6℃低下)だったのに対して、海水位低下のピークは約15,000年前(約130メートル低下)。
- 約6,000~5,000年前の縄文海進時は、意外にも温かさのピークではなく、10,000年前の方が温かかった。その時の海水位は約-30メートル。
ことが読み取れる。約2,000年の遅れについては、海洋大循環の2,000年周期と関係するのかも知れない。
なお、約10,000年前は現在よりも平均気温が2℃高かったので、現代では冷涼な興隆窪文化地域が快適だったのであろう。
「地質図 Navi」による 東・東南アジアの海面低下シミュレーション
産業総合研究所「地質図 Navi」の「海面上昇シミュレーション」は、名称から考えると、温暖化による海面の上昇で陸地がどれだけ水没するのかをシミュレートするのが本来の使い方らしい。しかし海水面が低下した時の陸地(大陸棚)を図示してくれる。
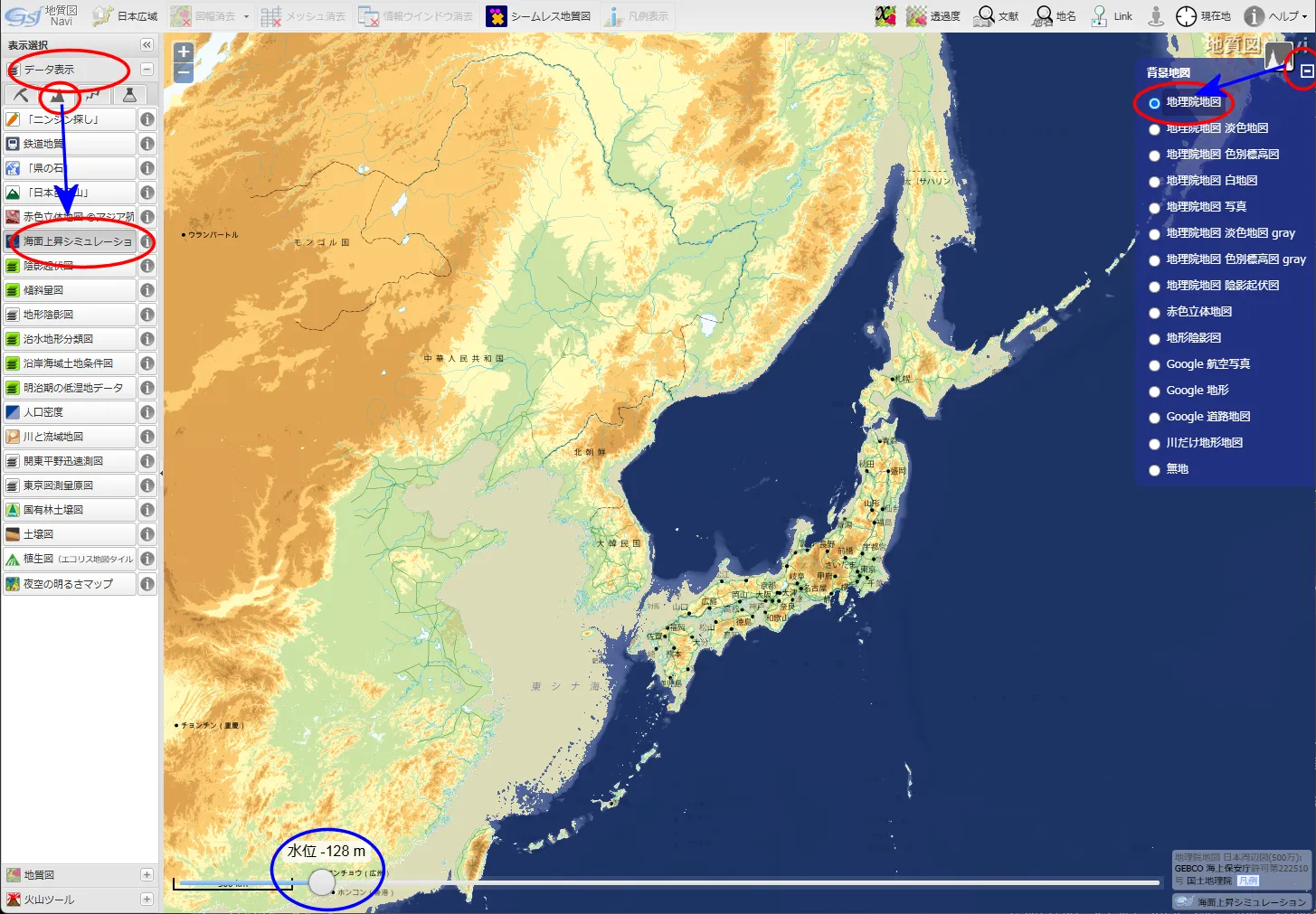
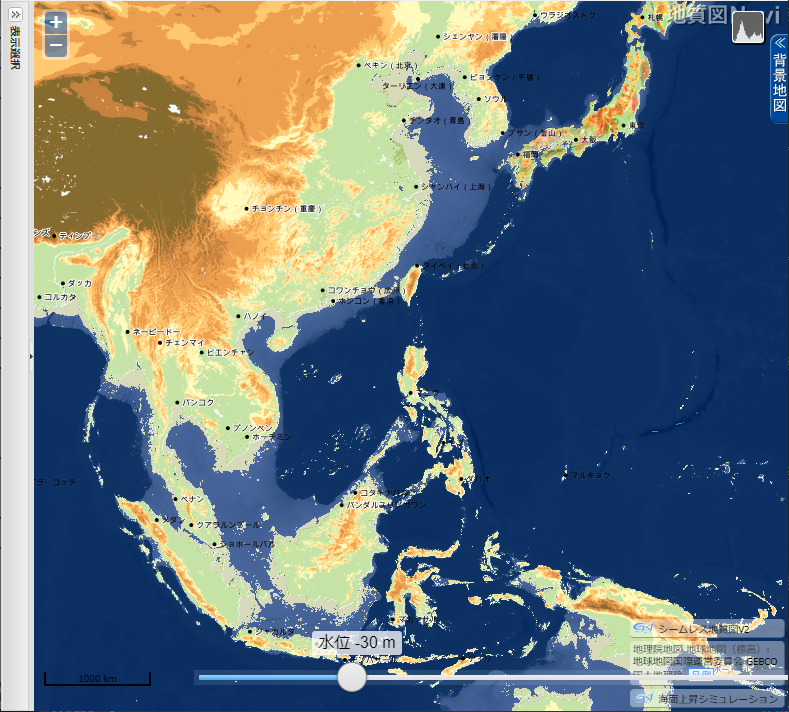
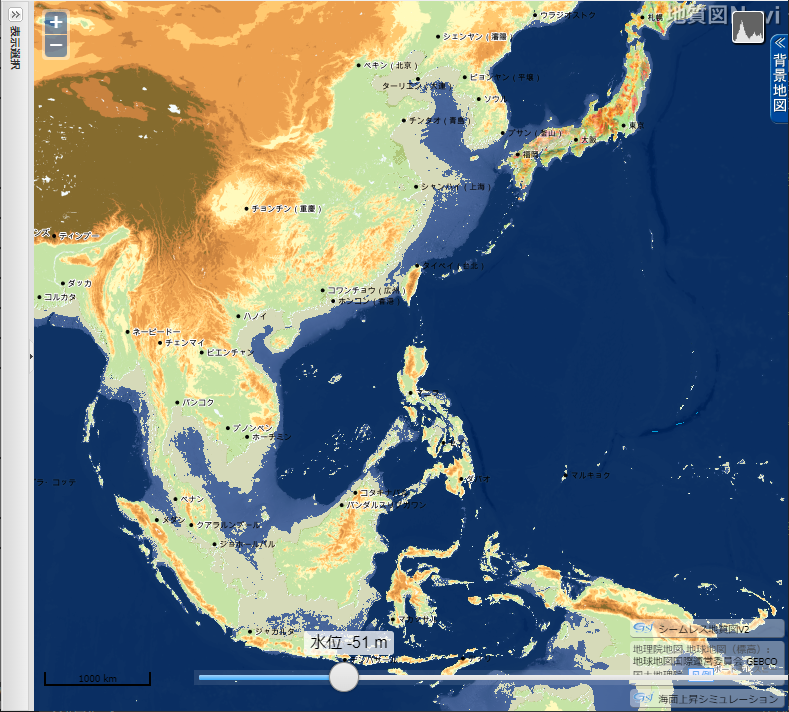
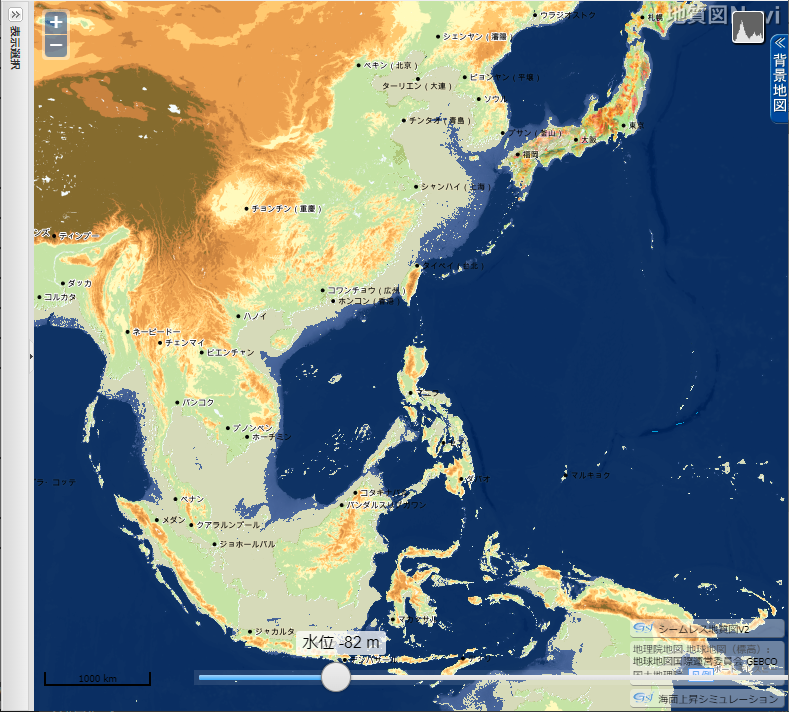
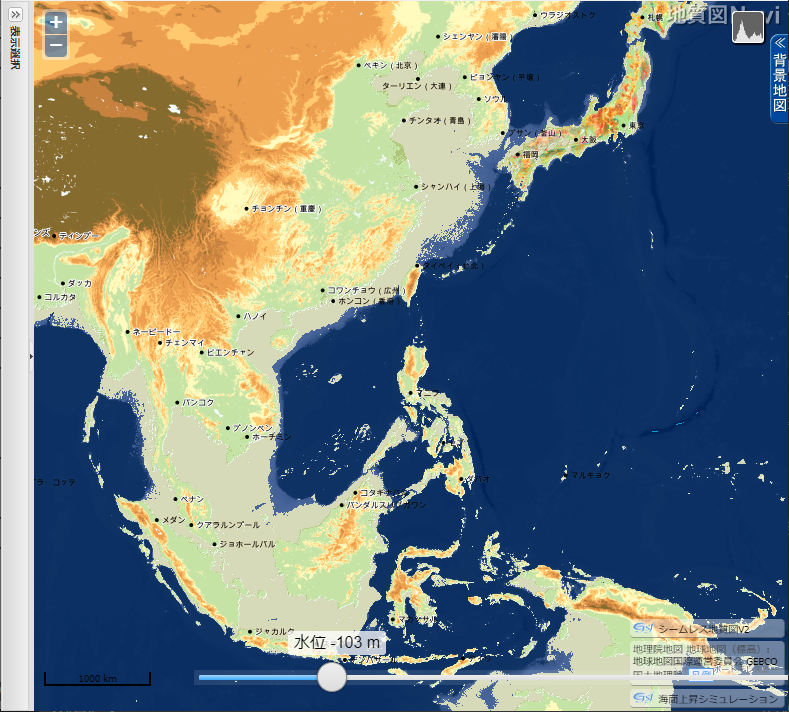
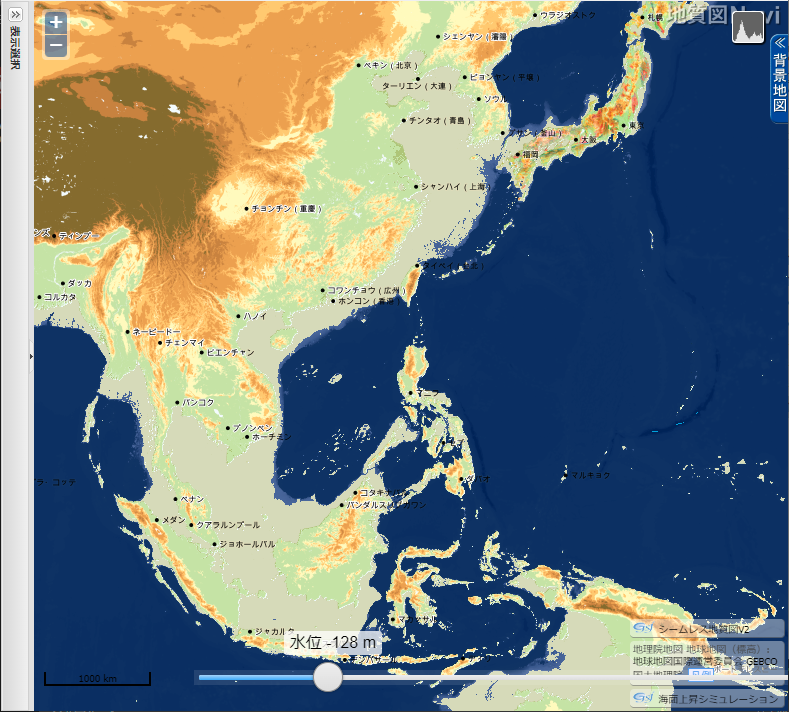
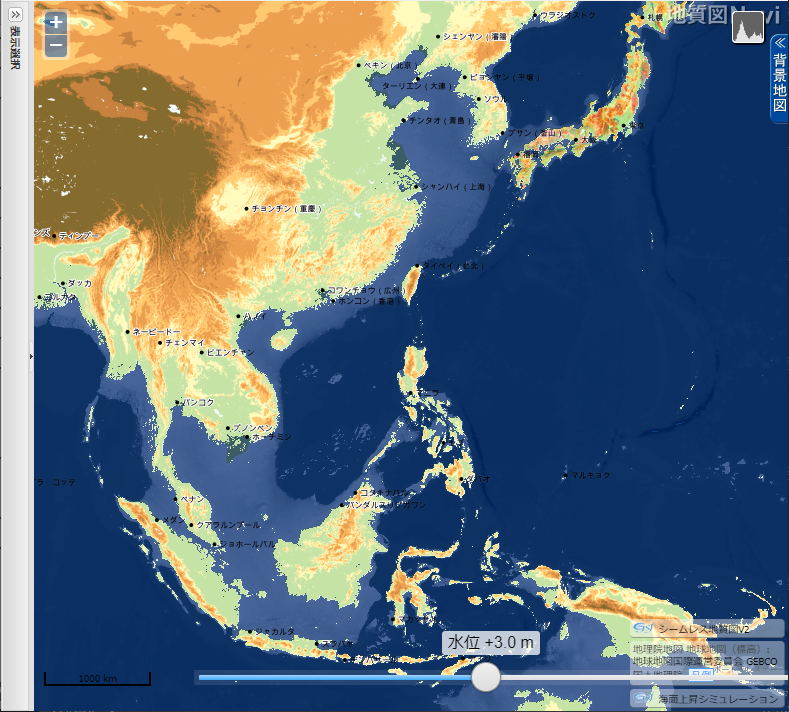
- 上の地図をクリック(タップ)すると拡大
最終氷期は、人類(ホモ・サピエンス)が出アフリカを果たした約7万年前に始まる。約33,000年前までは最悪の寒さではなく、海水位も-70~80 m の時期が長かったが、その後寒冷化が著しく進行して約25,000~16,000年前の最終氷期極大期(LGM=Last Glacial Maximun)には、平均気温が現在よりも5~6℃低く、海水面は120~130メートル低下したと推定されている。LGM の終了と縄文時代の開始が年代的に丁度重なる。
LGM の間、北アメリカとスカンジナビア半島を中心とするヨーロッパは、厚さ2,000~3,000メートルの氷床に覆われた無人の大地だったが、上の地図で示されるように南シナ海南部・タイランド湾の大陸棚は平原となっていた。それがスンダランドです。東シナ海の西側3分の2や台湾~ベトナムの東にも広大な平原があった。
氷河時代は、寒冷で厳しい気候というイメージがある。しかし当時の低緯度大陸棚は、現代よりも涼しかったので、現代の熱帯よりも快適だったでであろう。そのため人口急増 ➡ 食糧不足となって、日本列島のような中緯度への拡散を招いたのかも知れない。
これらの地図を比較すると......。
-
海水位 30 m 低下
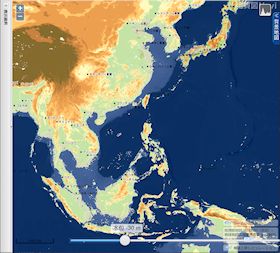
▲海水位 30m 低下 - 南シナ海には数多くの島が出現する。
- スマトラ島の東の平野がかなり広くなる。
- 台湾はまだ島である。
- 杭州湾が陸化し、北の渤海湾にかけて幅約 100 km の平野が出現する。
- 樺太島西の間宮海峡は既に陸化している(最深部約 20 m)。
-
海水位 51 m 低下
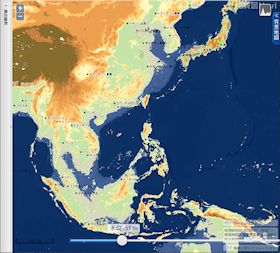
▲海水位 51m 低下 - スマトラ島とボルネオ島が地続きになる。
- インドシナ半島が南に張り出す。
- 上海の東で中国大陸が大きく東に張り出すようになる。その先端と五島列島の間は、約 250 km のみ。
- 台湾が大陸側と地続きかどうか微妙な状態になる。
- 瀬戸内海が大部分陸化する。
-
海水位 82 m 低下
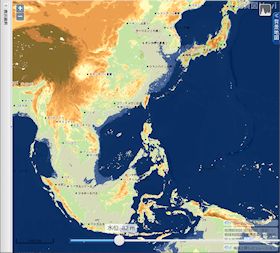
▲海水位 82m 低下(最終氷期の大部分) - 黄海の残り幅が僅かとなる。
- 現在の東シナ海の中央部にたくさんの小さな島々ができる。
- 東シナ海の西側半分が陸化する。
- 宗谷海峡が陸化する(最深部約 67 m)。
- 海水面の低下が 70~80 m で停滞していた時期が長かった(約3万年)ために形成された海岸段丘の化石がありそうである。
-
海水位 103 m 低下
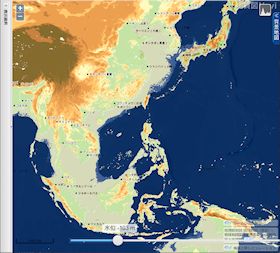
▲海水位 103m 低下 - 九州の西の大陸棚が陸化する。伊勢湾、豊後水道が消失し、隠岐の島が陸続きになる。屋久島・種子島はもう少しで大隅半島と繋がる。
- 朝鮮半島が大陸と一体化して半島でなくなる一方、済州島が半島になる。
- タイランド湾がほぼ陸化してスンダランドは最大面積近くになる(これ以上海水面が低下しても陸地面積はほとんど広がらない)。
-
海水位 128 m 低下
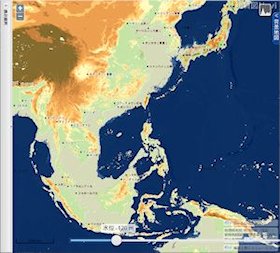
▲海水位 128m 低下(最終氷期極大期) - 現在の尖閣諸島の北にかなり大きな湾ができる。また、周辺は半島か島かわからないような地形となる。
- ニューギニア島とオーストラリア大陸の間が陸化してサフルランドとなる。
- 九州の北~西の大陸棚が陸化して、一体化した朝鮮半島・中国大陸と陸続きになりそうになる。
➥ -128m の関東~沖縄トラフ powered by「地質図 Navi」 - 現在の東シナ海の西側3分の2が陸化する。北ベトナムにかけても幅約 100 km の広大な平野が出現する。これらの土地は無人だったであろうか?
-
海水位 3 m 上昇
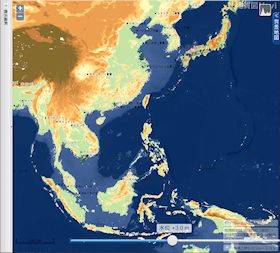
▲海水位 3m 上昇(縄文海進期) - 中国江蘇省北部のかなりの部分が水没する。
- 中国浙江省、福建省、広東省東岸の平野が更に狭くなる。
- 渤海湾沿岸の平野が狭くなる。
- 日本では霞ヶ浦周辺が入り江と化し、東京湾沿岸が水没して湾が広くなる。
陸地化したばかりの土地は森林にはならない。しばらくの間は草本類の天下でである。そんな亜熱帯から熱帯の陸地化して間もない環境(湿地や沼沢地)がイネ科の植物 Oryza rufipogon、即ち稲(Oryza sativa)の原種にとって好適だったのではないだろうか?
日本列島の黒曜石とサヌカイト主産地の分布
石器時代には石器の原材料となる黒曜石やサヌカイトが貴重品でした。白滝、隠岐の島、腰岳の黒曜石は、国内で使われただけでなく、朝鮮半島や沿海州、更には中国黒竜江省の松花江やモンゴルのアムール川(黒竜江)流域の遺跡でも出土する。
「Y染色体ハプログループの世界拡散を表す想定地図」を拡大すると、黒竜江省東部とバイカル湖南西に小さな D1a2(旧 D2)の分布域が見える。想定地図制作者は東向きの矢印を添えているが、逆に C 系統のように日本列島から石器(の材料?)を持って西進したのかも知れない。
良質な黒曜石を入手することは、金属器を持たない狩猟民族にとって死活問題だったであろう。
- アルファベットのマーカーは黒曜石、数字のマーカーはサヌカイト
関東~中部と九州北部を拡大 (開|閉)
サヌカイト主産地の分布
- 日本の黒曜石とサヌカイトの主産地の緯度・経度
-
- A. 白滝遺跡(黒曜石)43.874567,143.129350
北海道 遠軽町(旧白滝村) - B. 和田峠・霧ヶ峰周辺数ヶ所(黒曜石)36.109300,138.126100
長野県 下諏訪町東俣(東俣遺跡) - C. 恩馳島(黒曜石)34.186956,139.075992
東京都神津島村 ※神津島西隣の小さな島。神津島産よりも更に良質。 - D. 朝日・弁天山遺跡(黒曜石)35.224526,139.048610
神奈川県足柄下郡箱根町 - E. 皮子平(黒曜石)34.860244,138.979075
静岡県伊豆市筏場 - F. 柏峠遺跡(黒曜石)34.956300,139.063800
静岡県伊東市鎌田字落合 - G. 伊豆石丁場遺跡(黒曜石)35.053611,139.064444
静岡県熱海市上多賀 - H. 久見宮ノ尾遺跡(黒曜石)36.323892,133.231362
島根県隠岐の島町。同島南東部の「宮尾遺跡」と混同しないこと - I. 姫島(黒曜石)33.732889,131.642680
大分県東国東郡姫島村 - J. 腰岳(黒曜石)33.243044,129.870293
佐賀県伊万里市 - K. 牟田(黒曜石)33.373094,129.648664
長崎県佐世保市 横臼鼻近く - L. 東浜(黒曜石)33.132961,129.741664
長崎県佐世保市 淀姫神社 - M. 針尾中町(黒曜石)33.068567,129.752569
長崎県佐世保市 針尾送信所近く - N. 大崎半島(黒曜石)33.043627,129.826642
長崎県佐世保市川棚町 - 1. 峰一合遺跡(下呂石)35.801703,137.253326
岐阜県下呂市森 - 2. 二上山北麓(サヌカイト)34.544161,135.652717
奈良県・大阪府境 - 3. 金山(サヌカイト)34.305111,133.870923
香川県坂出市 - 4. 五色台(サヌカイト)34.344400,133.945860
香川県坂出市 ※サヌカイトの学名のもとになった讃岐石の産出地 - 5. 冠遺跡群(サヌカイト)34.435435,132.078904
広島・山口県境の冠高原 - 6. 鬼の鼻山北麓(サヌカイト)33.240577,130.097153
佐賀県多久市 - 7. 織島 岡本周辺(サヌカイト)33.324032,130.219236
佐賀県小城市
- 黒曜石の産地リストはウィキペディアの「黒曜石」の項目による。
- サヌカイトの産地リストはコトバンクの「サヌカイト」の項目による
- 数字は緯度と経度。少数点以下は10進法。場所によっては数 km の誤差を含むかも。
- A. 白滝遺跡(黒曜石)43.874567,143.129350
世界に目を向けてみると、ウィキペディア「黒曜石」の項目に掲載された国別黒曜石産地数の分布は次のようになっている。
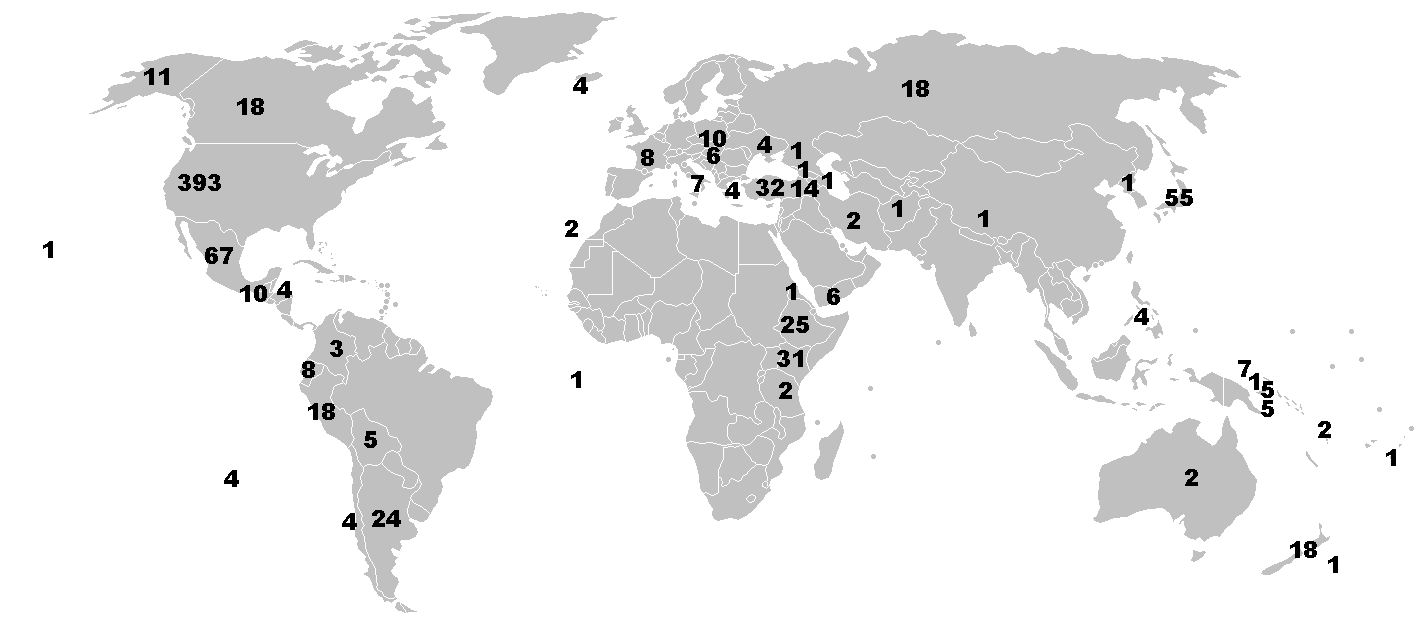
- クリック(タップ)すると拡大
- 数字は国ごとの産地数を示す。
- 出典:Kinori, CC BY-SA 3.0, via Wikimedia Commons
{kind=link}
米国が 393 地点とダントツに多く、次がメキシコの 67、日本は 55 地点で3位と結構健闘している。アジアでは文句なく1位なので、最終氷期の間陸化していた東シナ海、黄海を含めた東アジア全体への供給源だったのかも知れない。
蛇足であるが、「Wndy.com」という気象情報提供のサイトを紹介しておく。フル機能を使うには有料契約が必要であるが、無料でも海流を表示してもらえる。上のリンクを利用すれば海流が表示されるが、サイト検索からアクセスした場合、三本線(≡)メニューから表示情報として「海流」を選択すればよい。下の海上保安庁「海しる」の地図と比較してほしい。
西太平洋の海流
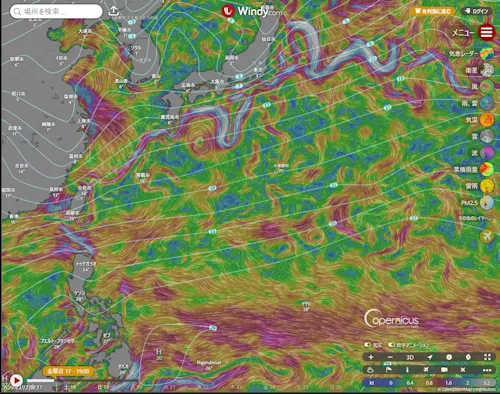
- 画像は「Windy.com」のスクリーンショット。画像をクリックすると Windy.com にジャンプ。
- 海上保安庁「海しる」の地図。
もしかすると、古代のある段階で、「ウナギの道」があったのかも知れない。十分な食糧と水を積んだ舟で宮古島あたりから黒潮に乗り、吐噶喇海峡から太平洋に抜けて本州沖を東上。伊豆諸島(恩馳島・神津島がある)で弱い海流を利用して南に向かい、マリアナ諸島で西向きの北赤道海流に乗ると、ウナギの産卵海域を通ってフィリピン・台湾沖経由で沖縄に戻れる。途中、もし小笠原諸島で西向きの海流(黒潮反流)に乗れれば、距離が短縮されるので好都合である。「福徳岡ノ場」海底火山の噴火で噴出した軽石が沖縄・奄美に漂着して大被害が出たのは記憶に新しい。嵐に遭わなければの話であるが......。
最終氷期~弥生時代の 東アジア
Y-DNAハプログループ分布(仮説)
最終氷期極大期の Y-DNAハプログループ分布地図を想像で作成してみた。
資料は、
- ハラストらによる Y-DNA 系統樹
- Maulucioni らのハプログループごとの分布図(D 系統、O 系統、N 系統、C2 系統など)
- 産総研のサイト「地質図 Navi」での「海面上昇シミュレーション」
である。残念ながら(当然ながら?)、私自身が原資料を作成したわけではない。また、あくまで最終氷期極大期(約 25,000年前~16,000年前)のものであって、極大期になるまでの約 35,000 年間や 16,000 年前以降を表現していない。
C1a2
- スマホ表示時、地図は左右スクロール可能。
- 産総研のサイト「地質図 Navi」の「海面上昇シミュレーション」で作成した地図を背景に楕円の推定分布域を重ねた。
Y-DNA 分布想像図
ストーリーとしては、次のようなものが考えてみた。ただし、あくまでたたき台となる仮説である。
- 人類(ホモ・サピエンス)は、東南アジアに遅れること数千年の約45,000年前に、ミャンマーから雲南経由、或いはインドシナ半島を横断して中国東部に到達した。北京近くの田園洞遺跡で発掘された約4万年前とされる田園洞人の Y-DNA ハプログループは K(L~T の祖型)。現在 K はオーストラリア大陸とニューギニアに分布する。
- 40,000~35,000年前に漁業と航海術を習得した C 系統が海沿いに日本列島に進出(海水面が 50~100 m 低下すると、東シナ海西部には小さな島が多数出現)。
- D 系統は「東シナ海平原」や「黄海平原」、現在の華北を中心に分布。西側が D1a1(後のチベット民族)、東が D1a2a(後の縄文系?)。
- 約29,000年前に姶良カルデラが破局噴火。九州の C 系統は壊滅、西日本も生態系に大打撃を受けた。C 系統は、数世紀の間大型獣に乏しく、狩猟民族には住みにくい環境になった西日本から、被害の少ない東日本に移住。
- 最終氷期極大期(25,000~16,000年前)、平均気温は現代よりも5~6℃低く、海水面の低下は最大 120~130 m に達した。日本海は津軽海峡と対馬海峡で太平洋に繋がっていたものの、水路が浅すぎて暖流である対馬海流が流入できなかったので、日本海で発生する水蒸気が少なく、日本列島は比較的乾燥していた。
- 最終氷期極大期を過ぎて温暖化が始まると N 系統と O 系統が北上して狭まりつつあった「東シナ海平原」と「黄海平原」に進出。しかし、約2,000年遅れで海水面の上昇が始まると、河川沿いに内陸に移動。
- D 系統は東西に分断され、D1a2a は日本列島西半分に進出。N 系統は大型獣を追って更に北方へ進出。
- 東日本の C 系統は温暖化による大型獣の減少を嫌って北上。C2 の一部はアムール川を遡上、一部はオホーツク海、ベーリンジア平原経由(もしかしたら千島列島、アリューシャン列島経由)でアメリカ大陸に進出。
-
上山遺跡の位置 - 地図をクリックすると Google Map で表示
- 約8,200年前に中国の遼河流域で N 系が遼河文明を起こした(興隆窪文化)。12,000年前頃から4,000年前頃までは豊かな水資源に恵まれ、湖沼や森林もある地域だったが、その後乾燥化した。その結果 N 系は次第に O2(漢民族)系や C2(モンゴル人など)に取って代わられた。現在の N 系統の分布から、ゾウやシカ類などの大型獣を追って北のアムール川流域や極北を含む北ユーラシアに移住したと考えられる。なお、4,000年前は黄河中下流域で夏が成立した時代である。
- 約7,300年前に鬼界カルデラが大噴火。南九州は再び壊滅、中・北九州~近畿の D1a2a は火山爆発が怖い西日本を捨てて東日本や朝鮮半島にしばらく避難。縄文時代の日本列島の人口が極端に東日本に偏っていたのは、この鬼界カルデラと前述の姶良カルデラの2回の大噴火が原因と思われる。
-
約4,000年前以降、華北で穀物栽培と畜産を組み合わせて黄河文明を起こし、強力になった漢民族(O2)に圧迫されて、D1a1(羌族≒チベット民族、彝族、納西族の先祖?)は西~南西に、O1 系各民族(苗・瑤語族、オーストロネシア語族)は南西~南東に移住開始。O1b2 は東~南東に移住して東夷となった。一部は漁民である C1a1 や日本列島の採集狩猟民である D1a2a と融合して倭人となったのだろう。
(以下 2025/07/06 追記)アルデヒド脱水素遺伝子2*504Lys(いわゆる「下戸遺伝子」)とも関係がありそうである。日本人は下戸遺伝子保有率が高いと言われるが、実は日本人よりも高いのが中国福建省人である。アルデヒド脱水素遺伝子2*504Lys
の分布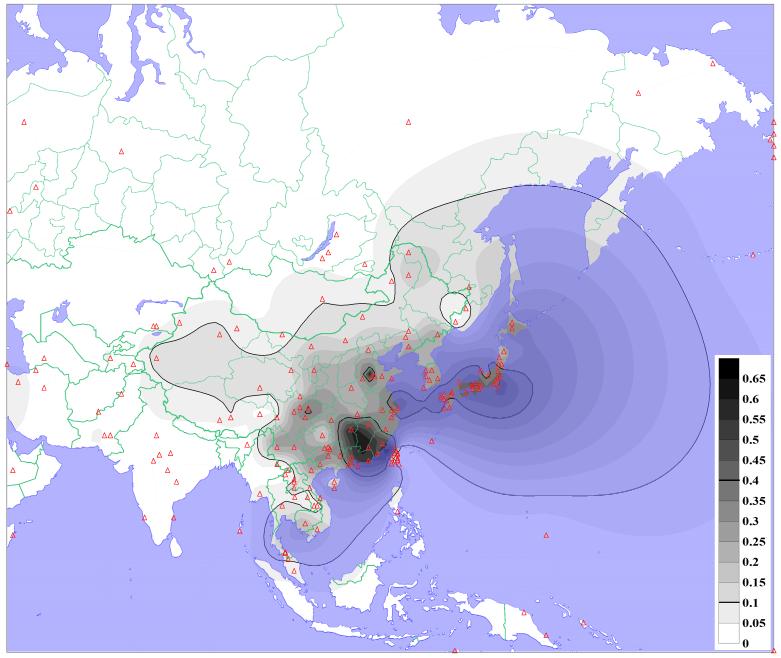
- 倭人は、C1a1 からの伝統である航海術を使って沖縄諸島に進出。中でも宮古島はタカラガイ(キイロダカラとハナビラダカラ)の養殖が行われていた。紐を通すための穿孔加工を施されたタカラガイは、沖縄本島に集積された後、東シナ海を左回りの航路で中国大陸に供給されて古代王朝の通貨となった。お金に関する漢字が貝偏をもつのはこのためである。(丸地三郎氏が熊本大学文学部木下尚子教授のタカラガイに関する研究を発展させている)
- 九州西岸、特に五島列島は、そばを対馬海流が流れる(➡ Windy.com の海流表示)ので、倭人の交易の中継基地となっていたに違いない。黒曜石の大産地である腰岳に近いことも好都合であった。佐賀県の有明海側にはサヌカイトの産地もある。
中国戦国時代、紀元前5世紀頃から紀元前3世紀にかけて、中国大陸東岸の国が相継いで滅亡した。
- 徐は紀元前 512 年に呉によって滅ぼされた。
- 呉は紀元前 473 年に越によって滅ぼされた。
- 越も紀元前 306 年に楚によって滅ぼされた。
- 楚も紀元前 223 年に秦によって滅ぼされた。
中国大陸では、滅ぼされた国の旧支配者階級は、うかうかしていると殺される運命となる。このため、朝鮮半島西岸にはこれらの国の遺民の社会ができていたのではないかと考えられる。
そうした状況の中で、徐国王の系譜に連なると思われる方士徐福が発見された。泰山で行う封禅儀式についての知見を持っている人はいないかと人材集めが行われたのかも知れない。伝承によると徐福は天文や航海術にも通じていたという。不思議なことに、徐福は童男童女3,000人、百工とともに蓬莱山にあるという不老不死の仙薬を探しに出航した。
不老長寿の霊薬探しというのは世間向けの口実で、徐福にとっては新天地が得られるかどうか、始皇帝にとっては徐福が拓いた新天地を支配できるかどうかという狐と狸の化かし合いだったのかも知れない。
- 紀元前 219 年、徐福第1回出航。開拓装備?
- 紀元前 210 年、連弩を装備して徐福が第2回出航。
2回目の出航にあたり、なぜ連弩を要望し、なぜその要望が叶えられたのか? 前述の丸地氏は、到着した倭=日本列島で在地勢力と戦いになってしまったのではないだろうかと推理している。
第2回出航後ほどなくして始皇帝の崩御すると動乱が起きて秦が滅び、紀元前 206 年に劉邦の漢(前漢)成立した。その結果、幸運なことに徐福捜索が行われなかった。
もし徐福伝説の通り、童男童女3,000人と百工を率いた徐福の一行が日本列島に到着していたら、新しい国を築く核になったことであろう(一人、或いは1家族で来ても埋没してしまう)。始皇帝からの追っ手の目を避けるために、玄界灘沿岸ではなく有明海側の湾の奥まった場所を本拠地とし、数十~数百 km 離れた数ヶ所に拠点を築くのは自然である。
ところで日本のコメは大きく分けて3種類ある。
- 熱帯ジャポニカ(焼畑)
- 温帯ジャポニカ(水田稲作)
- 極短粒米
- 短粒米
日本で発見された最も古いコメの遺物は、岡山県の縄文遺跡から出土した約6,000年前の米のプラントオパールであるが、焼畑で栽培された熱帯ジャポニカであるらしい。次の段階が、唐津市の菜畑遺跡など玄界灘沿岸から紀元前5世紀頃の水田稲作遺跡から出土した温帯ジャポニカの極短粒米である。しかしこれは有明海沿岸から広まった短粒種に取って代わられた。この短粒種をもたらしたのが徐福一行と考えられるのである。
なお、佐賀県や福岡県には大イチョウの名所がある。一方、徐福の出身地である琅琊郡には郯城(現在の行政区分は臨沂市郯城県)があり、特産品が銀杏である。郯城は古代徐国の根城であった。人類が誕生する前に日本では一旦絶滅したイチョウが、いつの間にか日本で復活したのは徐福一行のおかげであるのかも知れない。